補體調(diào)節(jié)分子的結(jié)構(gòu)及功
補體系統(tǒng)的激活為一種級聯(lián)反應(yīng)�����,但受到多種調(diào)節(jié)分子的嚴格控制�����,其反應(yīng)的程度和單一成分的反應(yīng)都是在生物反饋近代制下而進行的��,從而限制了活化的擴大化�,以維持補體水平的平衡。調(diào)節(jié)作用包括兩個方面��,即自身衷變失活及一些抑制物的滅活作用����。前者指已活化的補體分子均不穩(wěn)定,如不及時與靶細胞膜結(jié)合即迅速衰變失活���;后者是通過抑制物的作用而使已活化的分子失去活性����。這一節(jié)中僅涉及后一個方面����。
一、C1抑制物
C1抑制物(C1INH)是血清中高度糖基化的一種蛋白質(zhì)�,含糖量高達35-49%。*初由Ranoff和lepow(1957)所發(fā)現(xiàn)�,稱其為C1酯酶抑制劑,與引同時Schultze等則將其稱為α2神經(jīng)氨酸糖蛋白�。C1INH為一單鏈分子,由478個氨基酸殘基組成���,分子量為104kDa���,由478個氨基酸殘基組成,分子量為140kDa����,鏈內(nèi)有兩對二硫鍵(圖5-13)。C1INH調(diào)節(jié)的主要方式是���,與活化的C1r或C1s結(jié)合形成穩(wěn)定的復(fù)合物而導(dǎo)致C1絲氨酸蛋白酶失活�����。其作用機理是�,C1INH通過提供一個酷似C1r或C1s的正常底物的序列為“銹鉺”(“bait”)����,被c 1r或C1s裂解暴露出一個活性部位,然后再與C1r或C1s結(jié)合形成共價的酯鍵而發(fā)揮抑制作用�����。此外�,C1INH還可防止在缺乏抗體時��,C1以很低但仍有一定速率出現(xiàn)的自發(fā)激活���。正常情況下,血液中的大多數(shù)C1可被7倍于其克分子濃度的C1INH所結(jié)合�����,以防止C1由于構(gòu)象改變而引起的自發(fā)激活����。但C1同抗原、抗體復(fù)合物的結(jié)合�,可使C1從C1INH的抑制作用中而獲釋。除上述作用外��,C1INH還可抑制凝血因子Ⅻa���、Ⅺa�����、激肽釋放酶及纖溶酶���,因而其在凝血����、激肽和纖溶系統(tǒng)中也有重要的調(diào)節(jié)作用��。
經(jīng)對C1INH cDNA序列的分析發(fā)現(xiàn)�,C1INH與其它幾種絲氨酸蛋白酶抑制物(serpin)超家族成員(α1抗胰蛋白酶�����、α1抗糜蛋白酶及抗凝血酶Ⅲ等)約有30%的氨基酸同源性��,物別是在C端的120個氨基酸�。C1INH的編碼基因定位于第11號染色體的短臂11.2~長臂13亞區(qū)。C1INH先天性缺陷時�����,可導(dǎo)致遺傳性血管神經(jīng)性水腫(hereditary angioneurotic edema,HAE)��。近年報道應(yīng)用C1INH濃縮劑防治HAE效果良好�,但尚未廣泛用于臨床。
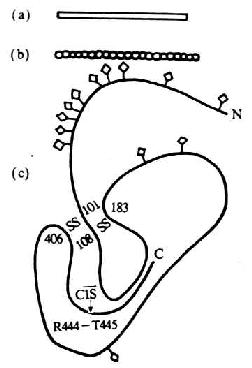
圖5-13 C1 INH分子的結(jié)構(gòu)模式圖
注:(a)為電鏡觀察模式圖
(b)為中子散射模式圖
(c)為園二色譜分析的二級結(jié)構(gòu)模式圖
二���、C4結(jié)合蛋白
C4結(jié)合蛋白(C4bp)是一種含量豐富的可溶性血清糖蛋白��,分子量為550kDa��,1977年由Ferreira等所報道�。其分子結(jié)構(gòu)模式現(xiàn)多以Dahlback等(1983)描述的“蜘蛛樣”(spiderlike)結(jié)構(gòu)來分析其結(jié)構(gòu)及功能。C4bp由8個亞單位組成��,電鏡下觀察形似蜘蛛��,其中有7條分子量相同(均為70kDa)的長鏈(α鏈)���,由549個氨基酸殘基組成���,鏈間以二硫鍵相連結(jié),并共同氨基酸殘基組成���,鏈間以二硫鍵相連結(jié)��,并共同連結(jié)于一中心體�����。α鏈含有8個SCR���,其N端是其頭部����,為與C4b相結(jié)合的部位���,可能位于第332-395位氨基本酸殘基��。第8條鏈(β鏈)較短(45kDa)由235個氨基酸殘基組成����,含有4個SCR為與蛋白S(PS)相結(jié)合的部位(圖5-14)��。C4bp以兩種方式抑制補體的活化�����。**種方式是�,通過其與C2競爭C4b��,而從c 4b2a中取代C2a��,并通過其與C4b的結(jié)合而陰止剩余的C2同C4b結(jié)合�����,由此抑制C3轉(zhuǎn)化酶的形成。C4bp與C4b的結(jié)合能力與細胞表面C4b分子的數(shù)成正比��,且較C2同C4b的結(jié)合能力高27倍��。**種方式是�����,C4bp作為I因子的一種輔助因子�����,促進I因子對C4b的裂解�����。有C4bp存在時��,I因子可將C4b的a`鏈完全裂解���;無C4pb時���,I因子的裂解作用不完全。其與I因子結(jié)合的活性部位位于第177-322位氨基酸殘基區(qū)域。PS與C4pb的第8條鏈(β鏈)結(jié)合����,不影響C4bp同C4b的結(jié)合,但可延長C4bp的半衷期��,從而強化C4bp的抑制作用����。
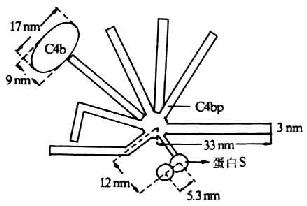
圖15-14 C4bp的結(jié)構(gòu)(模式圖)
C4bp基因定位于人的第1號染色體長臂32區(qū),同CR1��、CR2��、H因子��、MCP及DAF等的基因相連鎖���,并均含有數(shù)目不同的SCR。
三����、促衰變因子(CD55)
促衰變因子(decayaccelerating factor,DAF)是Nicholson-Weller等(1981)用正丁醇提取后,再以層析法從人和豚鼠紅細胞基質(zhì)中純化的一種膜蛋白��。因其具有促進C3轉(zhuǎn)化酶衷變的活性故名。經(jīng)在還原條件下做SDS-PAGE并以過碘酸-Schiff試劑染色表明����,純化的DAF為單鏈膜糖蛋白。人和豚鼠的DAF分了量不同分別為70kDa和60kDa?���,F(xiàn)已按白細胞分化抗原將其歸為CD55。Davitz等(1986)通過用磷脂酰肌醇(PI)特異性的磷脂酶C(PI-PLC)處理人外周血細胞可釋放DAF的事實探明��,DAF是經(jīng)糖磷脂酰肌醇(glycosylphodphatidylinositol,GPI)錨而固定于細胞膜中的�����。即糖蛋白的C末端共價結(jié)合于含PI的糖磷脂上����,再經(jīng)PI插入細胞膜脂質(zhì)雙層的外層小葉中。研究表明�,膜DAF的遷移率接近于膜類脂的遷移率,比大多數(shù)膜蛋白遷移率高一個數(shù)量級��。認為這有助于促進數(shù)目有限的膜DAF分子與細胞表面大量的C3b或C4b片段接觸�����。另外DAF的糖磷脂酰肌醇結(jié)構(gòu)還可能具有轉(zhuǎn)導(dǎo)細胞信號的作用。
除Nicholson-Weller等證實的分子量為70kDa的膜DAF外���,Kinoshita等(1987)用Western blotting在人紅細胞表面還檢出分子量為140kDa的一種膜DAF�,稱其為DAF-2��。DAF-2在膜上的數(shù)目不足70kDa膜DAF的1/10�����,但也有促進C3b轉(zhuǎn)化酶衰變的活性����,也含有GPI錨結(jié)構(gòu)。由于DAF-2的分子量較70kDa的膜DAF大一倍����,提示其為膜DAF的二聚體��,但用二巰基乙醇或以SDS使基變性���,都不能將DAF-2裂解成兩個成分�,故DAF-2的**結(jié)構(gòu)仍有待進一步闡明���。另外�����,近年應(yīng)用兩位點RIA測定法���,在血漿���、尿液、淚液���、唾液�、滑膜液和腦脊液���,以及組織培養(yǎng)上清中均檢出可溶性的DAF(sDAF)����,水平的40-400ng/ml范圍��。并發(fā)現(xiàn)尿液中的sDAF分子量略低于紅細胞上膜DAF的分子量�,疏水性也較膜DAF小,其抑制細胞表面C3轉(zhuǎn)化酶內(nèi)在裝配的活性較膜DAF約低100倍����,但仍具有促進已形成的C3轉(zhuǎn)化酶衰變的作用�,效能類似于C4bp�����。
膜DAF廣泛分布于各種血細胞及其他各處的細胞上�,包括紅細胞、粒細胞�����、單核細胞����、**細胞(T、B)���、血小板����、骨髓單個核細胞�、紅細胞的始祖細胞�����,角膜、結(jié)合膜����、消化道粘膜、外分泌腺����、腎小管、膀胱��、**粘膜����、胞膜、心包及滑膜的上皮細胞�����,精子����,以及培養(yǎng)的臍靜脈內(nèi)皮細胞上。但NK細胞上則缺如���。陣發(fā)性血紅蛋白尿(paroxysmal nocturnalhemohlobulinuria,PNH)病人的紅細胞上也缺少DAF��,并以缺乏程度將該病分為三個型��。PNH病人的紅細胞對補體介導(dǎo)的溶血作用高度敏感��,就是由于紅細胞上缺乏DAF及基它含GPI錨的分子而引起的����。DAF帶有Cromer抗原。極少數(shù)稱之為Inaba或IFC缺乏的Cromer相關(guān)抗原的個體也缺乏DAF��。
DAF生物學(xué)活性及生理功能已虱到充分證實�。它可保護宿主細胞免遭補體介導(dǎo)的溶解破壞。其作用機理是����,DAF不僅可阻止經(jīng)典或替代途徑C3和C5轉(zhuǎn)化酶的裝配,并且可通過誘導(dǎo)催化單位C2a或Bb的快速解離而使已形成的C4����、C5轉(zhuǎn)化酶失去穩(wěn)定性,從而抑制補體攻擊單位的活化(圖5-15)����。DAF的這種抑制作用**于直接結(jié)合在細胞上的C3���、C5轉(zhuǎn)化酶�����,也即DAF不抑制靶細胞上正常的補體激活劑如微生物和**復(fù)合物��。但DAF不能作為I因子裂解C3b和C4b的輔因子而發(fā)揮作用���。另外�����,DAF雖不能阻止C2和B因子(分別通過與C4b或C3b結(jié)合)與細胞的*初結(jié)合����,但卻可使C2a或Bb由它們結(jié)合的部位解離出來�����,以而阻止C3轉(zhuǎn)化酶的裝配���。
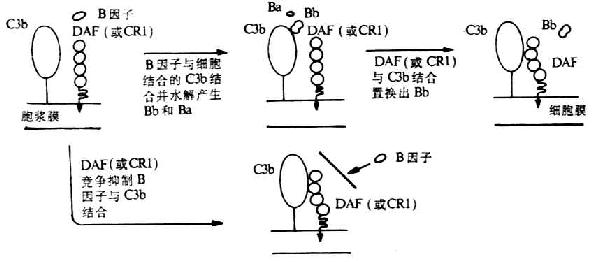
圖5-15 DAF抑制替代途徑中C3轉(zhuǎn)化酶形成的的機理
編碼人DAF的基因位于第1號染色體的長臂上32區(qū)一個800kb片段內(nèi)�����,與其它幾種補體激活調(diào)節(jié)劑(RCA)的基因緊密連鎖��,排列順序依次為:MCP-CR1-CR2-DAF-C4bp�����。其中DAF基因的長度約為C4bp�。其中DAF基因的長度約為35kb,以限制性酶譜分析表明為一單拷貝基因�。在DAF基因的非編碼區(qū)還有3個限制性酶切片段長度多態(tài)性(RFLP)結(jié)構(gòu),兩個為HindⅢ酶切位點���,1個為Bamh Ⅰ酶切位點����。DAF的cDNA已克隆成功��,并進行了核苷酸和氨基酸序列分析����。關(guān)于DAf cDNA和膜糖蛋白的結(jié)構(gòu)見圖5-16���。DAF的cDNA結(jié)構(gòu)從5`端開始依次為:信號肽區(qū)、四個SCR(長度1143bp)�����、富含絲氨酸與蘇氨酸(S/T)的區(qū)(約70個氨基酸)及疏水區(qū)���,*后終止于3`端的poly(A)。由cDNA推導(dǎo)的氨基酸序列得出DAF蛋白由381個氨基酸所組成��,包括34個氨基酸的信號肽�����,富含S/T的區(qū)為DAF中大多數(shù)延伸的O-連接的糖基化部位�����。SCR中有1上N-連接的的糖基化部位�。C末端的疏水區(qū)在翻譯后被糖磷脂所取代,為DAF分子與細胞膜相結(jié)合的部位����。
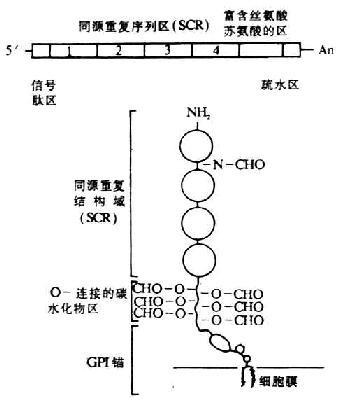
圖5-16 DAF cDNA和其膜糖蛋白的結(jié)構(gòu)
